Как происходит соединение аминокислот в белке. Понятие о ферментах. Строение белковых молекул
1. Классификация аминокислот.
2. Синтезы , , - аминокислот.
3. Свойства аминокислот: амфотерность, реакция по аминогруппе и карбоксилу.
4. - аминокислоты, их роль в природе.
5. Синтез пептидов.
Белковые вещества:
1. Классификация.
2. Строение. Первичная структура, понятие о вторичной, третичной и четвертичной структурах.
3. Понятие о ферментах.
Классификация аминокислот.
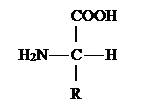
Аминокислотами называются органические кислоты, содержащие одну или несколько аминогрупп. В зависимости от природы кислотной функции аминокислоты подразделяют на аминокарбоновые, например H2 N(CH2)5 COOH, аминосульфоновые, например H2 N(CH2)2 SO3 H, аминофосфоновые, H2 NCH2, аминоарсиновые, например, H2 NC6 H4 AsO3 H2 .
Согласно правилам ИЮПАК название аминокислот производят от названия соответствующей кислоты; взаимное расположение в углеродной цепи карбоксильной и аминной групп обозначают обычно цифрами, в некоторых случаях - греческими буквами. Однако, как правило, пользуются тривиальными названиями аминокислот. (см. таблицу 1.).
В зависимости от положения аминогруппы по отношению к карбоксилу различают , и - аминокислоты:
Все - аминокислоты, кроме аминоуксусной (глицина), имеют асимметрический - углеродный атом и существуют в виде двух энантиомеров. За редким исключением, природные - аминокислоты относятся к L- ряду (S-конфигурация) и имеют следующее пространственное строение:
 По физическим и ряду химических свойств аминокислоты резко отличаются от соответствующих кислот и оснований. Они лучше растворяются в воде, чем в органических растворителях; хорошо кристаллизуются; имеют высокую плотность и исключительно высокие температуры плавления. Эти свойства указывают на взаимодействие аминных и кислотных групп, вследствие чего аминокислоты в твёрдом состоянии и в растворе (в широком интервале pH) находятся в цвиттер-ионной форме (т.е. как внутренние соли). Взаимное влияние групп особенно ярко проявляется у -
аминокислот, где обе группы находятся в непосредственной близости.
По физическим и ряду химических свойств аминокислоты резко отличаются от соответствующих кислот и оснований. Они лучше растворяются в воде, чем в органических растворителях; хорошо кристаллизуются; имеют высокую плотность и исключительно высокие температуры плавления. Эти свойства указывают на взаимодействие аминных и кислотных групп, вследствие чего аминокислоты в твёрдом состоянии и в растворе (в широком интервале pH) находятся в цвиттер-ионной форме (т.е. как внутренние соли). Взаимное влияние групп особенно ярко проявляется у -
аминокислот, где обе группы находятся в непосредственной близости.
 |
Цвиттер-ионная структура аминокислот подтверждается их большим дипольным моментом (не менее 50×10-30 Кл × м), а также полосой поглощения в ИК- спектре твердой аминокислоты или её раствора.
Таблица 1. Важнейшие аминокислоты.
| Тривиальное название | Сокр.название ос- татка ами нок-ты | Формула | Температура плавления, 0С. | Растворимость в воде при 250С, г/100г. |
| Моноаминомонокарбоновые кислоты | ||||
| Гликокол или глицин | Gly | H2 NCH2 COOH | 262 | 25 |
| Аланин | Ala | H2 NCH(CH3) COOH | 297 | 16,6 |
| Валин | Val | 315 | 8,85 | |
| Лейцин | Leu | 337 | 2,2 | |
| Изолейцин | He | CH3 ─ CH ─ C2 H5 | 284 | 4,12 |
| Фенилаланин | Phe | 283 (разл.) | ― | |
| Моноаминодикарбоновые кислоты и их амиды | ||||
| Аспарагиновая кислота | Asp(D) | 270 | 0,5 | |
| Аспарагин | Asn(N) | 236 | 2,5 | |
| Глутаминовая кислота | Glu(E) | 249 | 0,84 | |
| Глутамин | Gln(Q) | 185 | 4,2 | |
| Диаминомонокарбоновые кислоты | ||||
| Орнитин(+) | Orn | 140 | ― | |
| Лизин | Lys(K) | CH2 CH2 CH2 CH2 NH2 | 224 | Хорошо растворим |
| Аминокислоты | ||||
| Аргинин | Arg® | CH2 CH2 NH ─ C ─ NH2 | 238 | 15 |
| Гидроксиаминокислоты | ||||
| Серин | Ser(S) | 228 | 5 | |
| Треонин | Tre(T) | 253 | 20,5 | |
| Тирозин | Tyr(Y) | 344 | ― | |
| Тиоаминокислоты | ||||
| Метионин | Met(M) | 283 | 3,5 | |
 Цистин Цистин
| (Cys)2 | 260 | 0,011 | |
| Цистеин | Cys© | 178 | Хорошо растворим | |
| Гетероциклические аминокислоты | ||||
| Триптофан | Try(W) | 382 | 1,14 | |
| Пролин | Pro(P) | 299 | 16,2 | |
| Оксипролин | Opr | 270 | 36,1 | |
| Гистидин | His(H) | H2 C ―C ―― CH | 277 | 4,3 |
Синтезы , , - аминокислот.
- аминокислоты получают галогенированием карбоновых кислот или эфиров в -положение с последующей заменой галогена на аминогруппу при обработке амином, аммиаком или фталимидом калия (по Габриэлю).
По Штрекеру – Зелинскому - аминокислоты получают из альдегидов:
В этой реакции вместо альдегидов могут быть использованы кетоны, а вместо диалкилфосфитов- диалкилтиофосфиты, кислые эфиры алкилфосфонистых кислот RP(OH)OR и диарилфосфиноксиды Ar2 HPO.
Альдегиды и кетоны или их более активные производные – кетали служат для синтеза - аминокислот с увеличением числа углеродных атомов на две единицы. Для этого их конденсируют с циклическими производными аминоуксусной кислоты – азалактонами, гидантоинами, тиогидантоинами, 2,5-пиперазиндионами или с её медными или кобальтовыми хелатами, например:
 |
Удобные предшественники - аминокислот - аминомалоновый эфир и нитроуксусный эфир. К их - углеродным атомам можно предварительно ввести желаемые радикалы методами алкилирования или конденсации. - кетокислоты превращают в - аминокислоты гидрированием в присутствии NH3 или гидрированием их оксимов, гидразонов и фенилгидразонов.
 |
Можно получать - аминокислоты также непосредственно из - кетонокислот, действуя на них аммиаком и водородом над никелевым катализатором:
Некоторые L-- аминокислоты ввиду сложности синтеза и разделения оптических изомеров получают микробиологическим способом (лизин, триптофан, треонин) или выделяют из гидролизатов природных белковых продуктов (пролин, цистин, аргинин, гистидин).
Аминосульфоновые кислоты получают при обработке аммиаком продуктов присоединения NaHSO3 к альдегидам:
RCHO + NaHSO3 ® RCH(OH)SO3 Na ® RCH(NH2)SO3 Na
- аминокислоты синтезируют присоединением NH3 или аминов к , - ненасыщенным кислотам:В.М.Родионов предложил метод, в котором совмещаются в одной операции получение , -непредельной кислоты конденсацией альдегида с малоновой кислотой и присоединение аммиака:
 |
|
Аминокислоты получают гидролизом соответствующих лактамов, которые образуются в результате перегруппировки Бекмана из оксимов циклических кетонов под действием H2 SO4. -аминоэтановую и -аминоундекановую кислоты синтезируют из ,,,-тетрахлоралканов путем их гидролиза конц. H2 SO4 до -хлоралкановых кислот с последующим аммонолизом:
Cℓ(CH2 CH2)n CCℓ3 → Cℓ(CH2 CH2)n COOH → H2 N(CH2 CH2)n COOH
Исходные тетрахлоралканы получают теломеризацией этилена с CCℓ4.
Бекмановская перегруппировка оксимов циклических кетонов. Наибольшей практический интерес представляет перегруппировка оксима циклогексанона:
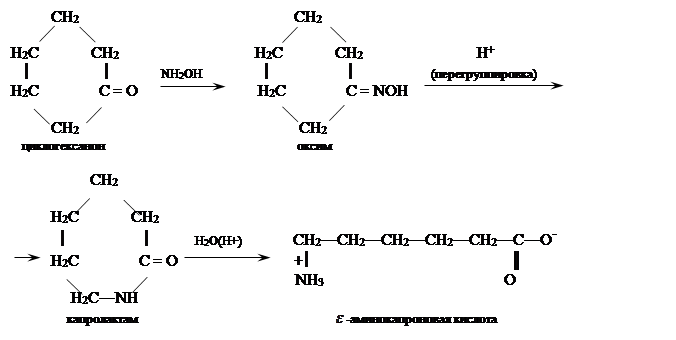
Получаемый этим путем капролактам полимеризуют в высокомолекулярный поликапромид
из которого изготовляют капроновое волокно.
Свойства аминокислот: амфотерность, реакция по аминогруппе и карбоксилу.
1. Большинство аминокислот – бесцветные кристаллические вещества, обычно хорошо растворимы в воде, часто сладковаты на вкус.
2. В молекулах аминокислот содержатся две группы с прямо противоположными свойствами: карбоксильная группа-кислотная, и аминогруппа с основными свойствами. Поэтому они обладают одновременно и кислотными и основными свойствами. Как кислоты, аминокислоты образуют со спиртами сложные эфиры, а с металлами и основаниями-соли:
Связь между атомом меди и азота осуществляется дополнительными валентностями(за счет свободной пары электронов азота аминогруппы). Как видно, при этом возникают кольчатые структуры, состоящие из пятичленных циклов. На легкость образования подобных пяти- и шестичленных циклов обратил внимание в 1906г. Л.А. Чугаев и отметил их значительную устойчивость. Медь(и другие металлы) в таких внутрикомплексных соединениях не имеют ионного характера. Водные растворы подобных соединений не проводят в заметной степени электрический ток.
При действии едких щелочей на медные соли аминокислот не происходит выпадания гидрата окиси меди. Однако при действии сероводорода происходит разрушение внутрикомплексного соединения и выпадает труднорастворимая в воде сернистая медью
3. Кислотные свойства в моноаминокислотах выражены весьма слобо-аминокислоты почти не изменяют окраски лакмуса. Таким образом, кислотные свойства карбоксила в них значительно ослаблены.
4. Как амины, аминокислоты образуют соли с кислотами, например:
HCℓ∙NH2 CH2 COOH
Но эти соли весьма непрочны и легко разлагаются. Таким образом, основные свойтва аминогруппы в аминокислотах также значительно ослаблены.
5. При действии азотистой кислоты на аминокислоты образуются оксикислоты:
NH2 CH2 COOH + NHO2 HOCH2 COOH + N2 + H2 O
Эта реакция совершенно аналогична реакции образования спиртов при действии азотистой кислоты на первичные амины.
6. С галоидангидритами кислот аминокислоты образуют вещества, которые одновременно являются и аминоксилотами и амидами кислот. Так, при действии хлористого ацетила на аминоуксусную кислоту образуется ацетиламиноуксусная килослота:
CH3 COСℓ + NH2 CH2 COOH СH2 CONHСH2 COOH + HCℓ
ацетиламиноуксусная килослота
Ацетиламиноуксусную кислоту можно рассматривать и как производное аминоуксусной кислоты, в молекуле которой атом водорода аминогруппы замещен ацетилом CH3 CO- и как ацетамид, в молекуле которого атом водорода аминогруппы замещен остатком уксусной кислоты -CH2 COOH.
7. - Аминокислоты принагревании легко отщепляют воду,
причем из двух молекул аминокислоты выделяются две молекулы воды и образуются дикетопиперазины:
Дикетопиперазины-циклические соединения, кольцо которых образовано четырьмя атомами углерода и двумя атомами азота. Дикетопиперазины - твердые, хорошо кристаллизующиеся вещества.
- Аминокслоты при нагревании теряют аммиак, переходя в непредельные кислоты:Лактамы можно рассматривать как внутренние амиды.
8. Аминокислоты образуют сложные эфиры при действии хлористого водорода на них спиртовые растворы. При этом, разумеется, образуется солянокислые соли эфиров, из которых свободные эфиры можно получить, удаляя хлористый водород окисью серебра, окисью свинца или триэтиламином:
NH3 -CH2 -C-O– + C2 H5 OH + HCℓ NH3 -CH2 -C-OC2 H5 Cℓ–
2 NH3 -CH2 -C-OC2 H5 Cℓ– + Ag2 O NH2 -CH2 -C-OC2 H5 + 2AgCℓ– + H2 O
Эфиры обычных аминокислот - жидкости, перегоняющиеся в вакууме. Именно этерификацией суммы аминокислот, получающихся в результате гидролиза белка, разгонкой в вакууме и последующим гидролизом Э.Фишер выделил индивидуальные аминокислоты и дал способ установления аминокислотного состава белков.
9. При действии пятихлористого фосфора на аминокислоты образуются солянокислые соли хлорангидридов аминокислот, довольно неустойчивые соединения, при отщеплении HCℓ образующие совсем неустойчивые свободные хлорангидриды:
NH3 -CH2 -C-O– + PCℓ5 NH3 -CH2 -C-Cℓ Cℓ– + POCℓ3
10. Аминокислоты ацилируются по аминогруппе:
NH2 -CH2 -C-O– + CH2 -C O CH3 -C-NH-CH2 -C-OH + CH3 -C-OH
║ ║ ║ ║ ║
ацетилгликокол
NH2 -CH2 -C-O– + C6 H5 -C-Сℓ C6 H5 -C-NH-CH2 -C-OH + HСℓ
бензоилгликокол
(гиппуровая кислота)
Образующаяся в последней из написанных реакций гиппуровая кислота – вещество, в виде которого травоядные животные выделяют с мочой небезвредную бензойную кислоту, попадающую в организм с пищей.
11. Аминокислоты можно алкилировать по аминогруппе. Алкилированием глицина получается метиламиноуксосная кислота-саркозин
NH3 -CH2 -C-O– + CH2 I CH3 NH2 -CH2 -C-O– + HI
саркозин
которая в связанном виде содержится в некоторых белках
При избытке иодистого метила образуется замещенная на четвертичноаммониевую группировку уксусная кислота
NH3 -CH2 -C-O– + 3CH3 I (CH3)3 N-CH2 -C-OH I– + 2HI
от которой можно отщепить HI и получить бетаин, лучше синтезируемый из триметиламина и хлоруксусной кислоты:
(СH3)3 N + CℓCH2 -C-ONa (CH3)3 N-CH2 -C-O– + NaCℓ
Бетаин, получивший свое название от свеклы(Betavulgaris), в соке которой он находится, дал название и всему классу внутренних солей, в которых анион и катион связаны внутри одной молекулы. В этом смысле говорят о бетаинобразной структуре самих аминокислот:
H3 N-CH2 -C-O– (CH3)3 N-CH2 -C-O–
Бетаины обладают большим дипольным моментом и солеобразны (тверды, нелетучи, водорастворимы).
- аминокислоты, их роль в природе.
В молекулах большинства - аминокислот содержится асимметрический атом углерода; природные аминокислоты существуют в виде оптических антиподов. Те из антиподов, конфигурация которых аналогична конфигурации правовращающего глицеринового альдегида, обозначаются буквой D; буквой L обозначаются антиподы, конфигурация которых соответствует конфигурации левовращающего глицеринового альдегида:
Конфигурация - асимметрического атома углерода у всех природных -аминокислот одинаковая, все они относятся к L-ряду.
Наибольший интерес представляют 20 - аминокислот, входящих в состав белковых молекул.
Принадлежность природных аминокислот к - аминокислотам легко устанавливается по их способности образовывать дикетопиперазины и характерные внутрикомплексные медные соли. Азотистая кислота превращает их в - оксикислоты, из которых многие известные уже со второй половины XVIII столетия. Если при этом получается неизвестная оксикислота, то аминогруппу можно заменить на хлор действием хлористого нитролиза
NH3 -CH-C-O– + NOCℓ Cℓ-CH-C-OH + N2 + H2 O
а затем, заменив действием цинка в кислой среде хлор на водород, получить известную карбоновую кислоту(в случае аминодикарбоновых кислот-двухосновную). Таким путем, например, из валина образуется изовалериановая кислота, из лейцина – изокапроновая, из глутаминовой – глутаровая, которые легко идентифицировать по каким-либо твердым производным (например по анилидам).
Синтез пептидов.
Дипептидами называются - амино-N-ациламинокислоты
R-CH-C-NH-CH-C-O–
Полипептиды построены по тому же амидному принципу из нескольких одинаковых или разных аминокислот. Они называются по числу участвующих остатков аминокислот ди-, три- и т.д. полипептидами.
Дипетиды с одинаковыми - аминокислотными остатками можно получить гидролитическим размыканием дикетопиперазинов. Дипетиды с любыми - аминокислотными остатками были получены Э. Фишером путем ацилирования аминокислоты по аминогруппе хлорангидритом - галоидзамещенной кислоты и последующей заменой - галоида на аминогруппу действием аммиака:
Сℓ-CH-C-Сℓ H2 N-CH-C-OH Сℓ-CH-C-NH-CH-C-OH + HСℓ
│║ │ ║│║ │ ║
ROR’ OROR’ O
Сℓ-CH-C-NH-CH-C-OH + 2NH3 H2 N-CH-C-NH-CH-C-O– + NH4 Сℓ
│║ │ ║ │║ │ ║
R O R’ O R O R’ O
Подобная же последовательность реакций, примененная к полученному дипетиту, приведет к трипетиду и т.д. Э. Фишер получил таким путем октадекапептид, состоящий из 18 остатков аминокислот.
В более новых методах синтеза полипептидов исходят из хлорангидридов аминокислот(или из иных функциональных производных аминокислот с резко выраженной ацилирующей способностью) с защищенной аминогруппой. Такая защита необходима, чтобы хлорангидрид первой аминокислоты не проацилировал себе подобную молекулу, а осуществил связь со второй аминокислотой. Защита аминогруппы ацетилированием мало удобна, так как условия удаления ацетильной группы гидролизом таковы, что сам ди или полипептид будет гидролизоваться, распадаясь на аминокислоты. Поэтому аминогруппу кислоты, предназначенной в качестве ацилирующего агента и превращаемой для этого в хлорангидрид, защищают, в водя в аминогруппу такую группировку, которую можно удалить из дипептида гидролизом в очень мягких условиях или каким-либо другим методом. Например группу CF3 CO- можно удалить обработкой слабой щелочью или гидрогенолизом; группу C6 H5 CH2 OCO- легко удалить гидрированием над палладиевым катализатором, восстановлением раствора натрия в жидком аммиаке или действием гидрозина; фталильная группа под действием гидразина отщепляется в виде
Что касается того, в форме какого функционального производного должен находится карбоксил защищенной описанным способом аминокислоты, то чаще чем хлорангидриды, применяют легко ацитилирующие эфиры или смешанные ангидриды, например:
HN-CH-C-O- -NO2 или HN-CH-C- -O-C-OR’
│ │ ║ │ │ ║ ║
где X- защищающий аминогруппу заместитель, R’- остаток пространственно затрудненного алифатического спирта, например (CH3)2 CH-CH2 OH, что обеспечивает разрыв ацилирующей молекулы по линии, намеченной пунктиром.
Совершенно иное дело-получение полипептидов, даже с высоким молекулярным весом, из остатков одной кислоты. Для этой цели выработан следующий метод(Лейхс), рассмотренный на примере глицина (R=H):
C6 H5 CH2 -O-C-Cℓ + H2 N-CH-C-OH
C6 H5 CH2 -O-C-NH-CH-C-OH
│ нагревание
C6 H5 CH2 -O-C-NH-CH-C-Cℓ O
Такие циклические внутренние смешанные ангидриды при нагревании распадаются с выделением CO2 и образованием высокомолекулярных полипетидов:
n O nCO2 + -HN-CH-C-(HN-CH-C-)n-1
В химии белков и полипептидов для сокращения принято писать формулы, обозначая остатки аминокислот буквами. Например, Glu- обозначение глутаминовой кислоты, Cys- цистеина, Gly- глицина и т.д. Сокращенная формула глутатиона будет в таком изображении
Белковые вещества. Классификация.
Белки ― высокомолекулярные природные полимеры, построенные из остатков аминокислот, соединенных амидной (пептидной) связью ―CO―NH―.
Каждый Б. характеризуется специфической аминокислотной последовательностью и индивидуальной пространственной структурой (конформацией). На долю белков приходится не менее 50% сухой массы органических соединений животной клетки. Функционирование белка лежит в основе важнейших процессов жизнедеятельности организма. Обмен веществ (пищеварение, дыхание и др.), мышечное сокращение, нервная проводимость и жизнь клетки в целом неразрывно связаны с активностью ферментов - высокоспецифичных катализаторов биохимических реакций, являющихся белками. Основу костной и соединительной тканей, шерсти, роговых образований составляют структурные белки. Они же формируют остов клеточных органелл (митохондрий, мембран и др.). Расхождение хромосом при делении клетки, движение жгутиков, работа мышц животных и человека осуществляются по единому механизму при посредстве белка сократительной системы (напр., Актин, Миозин). Важную группу составляют регуляторные белки, контролирующие биосинтез белка и нуклеиновых кислот. К регуляторным белкам относятся также пептидно-белковые гормоны, которые секретируются эндокринными железами. Информация о состоянии внешней среды, различные регуляторные сигналы (в т. ч. гормональные) воспринимаются клеткой с помощью спец. рецепторных белков, располагающихся на наружной поверхности плазматической мембраны. Эти белки играют важную роль в передаче нервного возбуждения и в ориентированном движении клетки (хемотаксисе). В активном транспорте ионов, липидов, сахаров и аминокислот через биологические мембраны участвуют транспортные белки, или белки-переносчики. К последним относятся также гемоглобин и миоглобин, осуществляющие перенос кислорода. Преобразование и утилизация энергии, поступающей в организм с питанием, а также энергии солнечного излучения происходят при участии белков биоэнергетической системы (напр., родопсин, цитохромы). Большое значение имеют пищевые и запасные белки (напр., Казеин, Проламины), играющие важную роль в развитии и функционировании организмов. Защитные системы высших организмов формируются защитными белками, к которым относятся иммуноглобулины (ответственны за иммунитет), белки комплемента (ответственны за лизис чужеродных клеток и активацию иммунологической функции), белки системы свертывания крови (напр. Тромбин, Фибрин) и противовирусный белок интерферон.
По составу белки делят на простые, состоящие только из аминокислотных остатков (протеины), и сложные (протеиды). Сложные могут включать ионы металла (металлопротеиды) или пигмент (хромопротеиды), образовывать прочные комплексы с липидами (липопротеины), нуклеиновыми кислотами (нуклеопротеиды), а также ковалентно связывать остаток фосфорной кислоты (фосфопротеиды), углевода (гликопротеины) или нуклеиновой кислоты (геномы некоторых вирусов).
По ряду характерных свойств протеины можно разделить на несколько подгрупп:
Альбумины. Они растворимы в воде, свёртываются при нагревании, нейтральны, сравнительно трудно осаждаются растворами солей. Примерами их могут служить: альбумин белка куриного яйца, альбумин кровяной сыворотки, альбумин мускульной ткани, молочный альбумин.
Глобулины. Они нерастворимы в воде, но растворяются в очень слабых растворах солей. Более концентрированными растворами солей они вновь осаждаются; осаждение происходит при меньшей концентрации, чем та, которая необходима для осаждения альбуминов. Эти белки являются очень слабыми кислотами. Примерами глобулинов могут служить: фибриноген, глобулин кровяной сыворотки, глобулин мускульной ткани, глобулин белка куриного яйца.
Гистоны. Белки основного характера. Находятся в виде нуклеопротеидов в лейкоцитах и красных кровяных шариках.
Протамины. Не содержат серы, обладают сравнительно сильными основными свойствами, дают кристаллические соли; содержатся (в виде нуклеопротеинов) в сперматозоидах рыб.
Проламины. Находятся в зернах различных хлебных злаков. Замечательной их особенностью является растворимость в 80% -ном спирте. Представителем этих белков может служить глиадин, составляющий главную часть клейковины.
Склеропротеины. Нерастворимые белки, которые составляют наружный покров тела животного и находятся в скелете и в соединительной ткани. К ним относятся кератин, коллагены, эластин, фиброин.
Керотин является главной составной частью волос, рогов, копыт, ногтей, перьев и верхнего слоя кожи. Скорлупа куриного яйца состоит из извести и кератина. Если растворить известь скорлупы яйца в кислоте, то останется мягкая кожа, состоящая из кератина; из кератина состоит кожица, которая следует за скорлупой яйца. По химическому составу кератин богат серой.
Коллагены. Чрезвычайно распространены в живых организмах. Из коллагенов состоит соединительная ткань; они находятся в хрящах. Кости позвоночных животных состоят из неорганических веществ (фосфорнокислого и углекислого кальция), жира и коллагенов.
При кипячении с водой или при действии перегретого водяного пара коллагены образуют клей. Если из костей извлечь жир и потом, обработав их кислотой, растворить фосфорнокислый кальций, то останется белковое вещество-оссеин. При обработке оссеина перегретым водяным паром он переходит в клей. Чистый костяной клей называется желатиной. Особенно чистая желатина получается из рыбьего пузыря кипячением с водой.
Эластин входит в состав жил и других эластичных веществ соединительной ткани.
Нити сырого шелка состоят из белкового вещества-фиброина, покрытого другим белковым веществом, играющим роль шелкового клея,-серицином. При кипячении с водой шелк освобождается от клея который при этом переходит в раствор.
Протеиды также можно разделить на несколько групп: фосфоропротеиды содержат в своем составе фосфор. Они, в противоположность протаминам, обладающим, как указано выше, основными свойствами, имеют определенно выраженный кислотный характер.
Главнейшим представителем фосфоропротеидов является казеин молока. Он обладает настолько ясно выраженным кислотным характером, что разлагает углекислые соли с выделением углекислого газа. Казеин растворяется в слабых растворах щелочей, образуя с ними соли. Соли казеина называются казеинатами.
При нагревании казеин не свертывается. При действии кислот на соли казеина он выделяется в свободном виде. Этим объясняется свертывание молока при прокисании.
Казеин применяется для изготовления твердой, напоминающей рог пластмассы-галалита).Для получения галалита казеин смешивают с водой, красками и наполнителями, прессуют под давлением, и полученные пластины обрабатывают формалином. Казеин содержит фосфор в виде сложного эфира фосфорной кислоты.
Из других фосфоропротеинов следует отметить вителлин, который находится в желтке куриного яйца.
Нуклеопротеиды находятся в клеточных ядрах. При осторожном гидролизе они расщепляются на белок и нуклеиновую кислоту.
Нуклеиновые кислоты являются весьма сложными веществами, расщепляющимися при гидролизе на фосфорную кислоту, углеводы и азотосодержащие органические вещества группы пиримидина и группы пурина.
Хромопротеиды. Под этим названием известны протеиды, которые представляют собой сочетание белков с окрашенными веществами. Из хромопротеидов наиболее изучен гемоглобин красящее вещество красных кровяных шариков. Гемоглобин, соединяясь с кислородом,
превращается в оксигемоглобин, который, отдавая свой кислород другим веществам, снова превращается в гемоглобин. Значение гемоглобина в жизни человека и животных очень велико. Он играет роль переносчика кислорода от легких к тканям. Образовавшийся в легких оксигемоглабин кровью разносится по телу и, отдавая свой кислород, способствует протекание в организме окислительных процессов. Кроме того, гемоглобин вместе с плазмой крови осуществляет регуляцию величины pH крови и перенос углекислоты в организме.
Характерной особенностью гемоглобина является его способность соединятся с окисью углерода, после чего он теряет способность соединяться с кислородом. Этим объясняется ядовитое действии окиси углерода.
Гемоглобин представляет собой соединение белка глобина с красящим началом гемохромогеном. Вне организма гемоглобин, при действии воздуха, превращается в метгемоглобин, который отличается от оксигемоглобына прочностью связи с кислородом. При обработке ледяной уксусной кислотой метгемоглобин расщепляется с образованием глобина и гематина C34 H32 O4 N4 Fe(OH). Обработкой метгемоглобина тем же реактивом, но в присутствии NaCℓ, получается хлористая соль гематина, называемая гемином, C34 H32 O4 N4 FeCℓ. Гемин образует характерные красно-коричневые таблички, которые дают возможность открыть присутствие крови в пятнах даже через несколько лет. Гематин очень близок к гемохромогену, но все же от него отличается.
Глюкопротеиды. Некоторые белки этой группы встречаются в слизистых соединениях животных организмов и обусловливаются свойства этих выделений тянуться в нити даже при сравнительно большом разбавлении. Эти белки образуются в подчелюстной железе(подчелюстная железа-одна из слюнных желез), печени, железах желудка и кишечника. Другие глюкопротеиды находятся в хрящах, яичном белке, стекловидном теле глаза и т.д. Исследованныепредставители глюкопротеидов являются сочетанием белков с веществами, содержащими остатки некоторых производных углеводов, серной и уксусной кислот.
Строение белковых молекул.
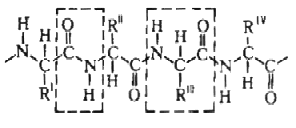
Практически все белки построены из 20 -аминокислот, принадлежащих, за исключением глицина, к L-ряду. Аминокислоты соединены между собой пептидными связями, образованными карбоксильной и -аминогруппами соседних аминокислотных остатков Белковая молекула может состоять из одной или нескольких цепей, содержащих от 50 до нескольких сотен (иногда –более тысячи) аминокислотных остатков. Молекулы, содержащие менее 50 остатков часто относят к пептидам. В состав многих молекул входят остатки цистина, дисульфидные связи которых ковалентно связывают участки одной или нескольких цепей. В нативном состоянии макромолекулы белка обладают специфичной конформацией. Характерная для данного белка конформация определяется последовательностью аминокислотных остатков и стабилизируется водородными связями между пептидными и боковыми группами аминокислотных остатков, а также гидрофобными и электростатическими взаимодействиями.
Различают четыре уровня организации белковых молекул.
Первичной структурой называют последовательность аминокислотных остатков в полипептидной цепи. Все белки различаются по первичной структуре, потенциально их возможное число практически неограничено.
Вторичная структура
белка – это -спираль, которая образуется в результате скручивания полипептидной цепи за счет водородных связей между группами: ―C― и ―N―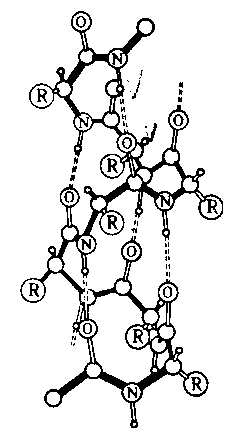 .
.
В одном витке спирали обычно содержится 3,6 аминокислотных остатка, шаг спирали – 0,544 нм.
Под третичной структурой белка понимают расположение его полипептидной цепи в пространстве. Существенное влияние на формирование третичной структуры оказывают размер, форма и полярность аминокислотных остатков. Третичная структура многих белков составляется из нескольких компактных глобул, называемых доменами. Между собой домены обычно бывают связаны тонкими перемычками – вытянутыми полипептидными цепями.
Термин четвертичная структура относится к макромолекулам, в состав которых входит несколько полипептидных цепей (субъединиц), не связанных между собой ковалентно. Между собой эти субъединицы соединяются водородными, ионными, гидрофобными и другими связями. Примером может служить макромолекула гемоглобина.
![]()
Понятие о ферментах.
Катализ многочисленных биохимических реакций, протекающих в растительных и животных организмах и их клетках, так же как и в одноклеточных микроорганизмах, совершается ферментами (энзимами).
Ферменты представляют, собой вещества или чисто белковые структуры, или протеиды-белки, связанные с небелковой простетической группой. Число уже известных ферментов очень велико. Считают, что одна клетка бактерии использует до 1000 разных ферментов. Однако лишь для не многих установлено строение. Примерами чисто белковых ферментов могут служить протеолитические ферменты пищеварения, такие как пепсин и трипсин. Известны случаи, когда один и тот же белок несет в организме и структурную и ферментную функцию. Примером служит белок мышц миозин, каталитически разлагающий аденозинтрифосфат – реакция, в данном случае дающая энергию сокращения мышцы.
Витамины – группы веществ обычно довольно сложной структуры, часто очень далеких химически друг от друга и объединяемых только по биологическому признаку. Это – требуемые в микроколичествах, но совершенно необходимые составные части пищи, недостаток которых вызывает болезнь, а отсутствие – гибель организма. Используя витамины(но не только их), организм человека и животных и строит свои ферменты. Одно и тоже вещество, служащее для организмов одного вида витамином, для других организмов может и не быть витамином по двум причинам: либо он может его синтезировать сам. Так аскорбиновая кислота – витамин для человека, но для крысы она не является витамином, так как организм крысы создает ее из глюкозы. Некоторые витамины имеют весьма универсальный характер и необходимы всем организмам – от одноклеточных(дрожжей и бактерий) до человека. Зеленые растения способны производить все свои вещества из минеральных исходных веществ и CO2 и, следовательно, не нуждаются в витаминах.
Для действия ряда белковых ферментов в организме необходимо присутствие микроколичеств ионов некоторых металлов-активаторов (Mg, Zn, Mo, Mn, Cu), которые витаминами не считаются. В состав некоторых коферментов и соответствующих витаминов входят металлы (железо, кобальт). Очень существенна роль микроколичеств металлов (микроэлементов) для развития растений; по-видимому, эти микроэлементы используются для постороения ферментов и служат как бы «витаминами растений».
Ферменты являются катализатороми, но отличаются от обычных катализаторов. Так им свойственны:
1. Гораздо большая специфичность по отношению к структуре катализируемого объекта и по отношению к реакции.
2. Полная стереохимическая специфичность.
3. Гораздо большая скорость протекания ферментных реакций по сравнению с теми же реакциями, катализируемыми обычнами катализаторами.
4. Невысокие оптимумы температуры их действия(обычно активность ферментов резко падает к 50О С и в интервале 50-100О С действие их полностью прекращается).
5. Денатурируемость- необратимая потеря каталитической активности при нагревании до 50-100О С. Существуют, однако, ферменты (трипсин, рибонуклеаза), активность которых восстанавливается по охлаждении даже после кипячения.
6. Существование оптимума кислотной среды для действия каждого фермента.
Высокая скорость ферментативных реакций объясняется, с одной стороны, как всегда при катализе, сильным снижением энергии активации реакции. Так, при катализе, сильным снижением энергии активации реакции. Так, при гидролизе казеина кислотой энергии активации 20,6 ккал/моль, а при гидролизе трипсином-только 12 ккал/моль. Гидролиз сахарозы кислотой требует энергии активации 25,5 ккал/моль, а ферментативный (сахарозой)-лишь 12-13 ккал/моль. С другой стороны, в ферментативных реакциях не меньшую роль играет предэкспоненциальный множитель уравнения Арреннуса, так как величина этого множителя, как правило, на много порядков выше, чем в реакциях обычного типа. Есть доказательство того, что ферменты содержат центры(«карманы»), фиксирующие субстрат на поверхности их молекул, и вторые центры, осуществляющие реакцию. Фермент может быть активен в том смысле, что он подтягивает активный центр к месту действия, несколько изменяя свою вторичную или третичную структуру.
Список используемой литературы:
1. А.Н.Несмеянов, Н.А.Несмеянов «Начала органической химии», т.1, 2.
2. Б.А.Павлов, А.П.Терентьев «Курс органической химии».
3. Химическая энциклопедия, т. 1.
4. Дж.Робертс, М.Касерио «Основы органической химии».
5. Шабаров Ю.С. «Органическая химия»
Саратовский Государственный Университет им. Н.Г.Чернышевского.
АМИНОКИСЛОТЫ
Аминокислоты – это строительные блоки макромолекул – белков. По строению
они являются органическими карбоновыми кислотами, у которых, как минимум, один
атом водорода замещен на аминогруппу.
КЛАССИФИКАЦИЯ
1. По абсолютной конфигурации мо-
лекулы – D- и L-формы.
Различия связаны со взаимным
расположением четырех замещающих
групп, находящихся в вершинах воображаемого тетраэдра. Как известно,
тетраэдр – это довольно жесткая
структура, в которой невозможно произвольным образом передвинуть вершины. Точно так
же для молекул, построенных на основе атома углерода. За эталон конфигурации принята структура молекулы глицеральдегида, установленная с помощью рентгеноструктурного анализа. Принято, что в качестве маркера используют наиболее сильно окисленный атом углерода, связанный с асимметричным атомом углерода (на схемах его располагают сверху). Таким атомом в молекуле глицеральдегида служит альдегидная группа, для аланина маркерной является СООН-группа. Атом водорода располагают так же, как
в глицеральдегиде.
В дентине, белке зубной эмали, скорость рацемизации L-аспартата равна 0,10% в год. При формировании зуба у детей используется только L-аспартат. Это позволяет определять возраст долгожителей в сомнительных случаях. Для ископаемых ос-
танков наряду с радиоизотопным методом также используют оп-
ределение рацемизации аминокислот в белке.
В белке любого организма содержится только один изомер, для млекопитающих это L-аминокислоты. Однако оптические изомеры претерпевают самопроизвольную неферментативную рацемизацию, т.е. L-форма пе-
реходит в D-форму. Это обстоятельство используется для
определения возраста ткани.
2. В зависимости от положения аминогруппы
Выделяют α, β, γ и другие аминокислоты. Для организ-
ма млекопитающих наиболее характерны -аминокислоты. 3.По оптической активности – право- и левовращаю-
Наличие ассиметричного атома С (хи-
рального центра) делает возможным только два расположения химических групп вокруг него. Это приводит к особому отличию веществ друг от друга, а именно – изменению
направления вращения плоскости поляриза-
ции поляризованного света, проходящего через раствор. Величину угла поворота
определяют при помощи поляриметра. В соответствии с углом поворота выделяют
правовращающие (+) и левовращающие (–) изомеры.
Деление на L- и D-формы не соответствует делению на право- и левовращаю-
щие. Для одних аминокислот L-формы (или D-формы) являются правовращающими,
для других – левовращающими. Например, L-аланин – правовращающий, а L-фенилаланин – левовращающий. При смешивании L- и D-форм одной аминокис-
лоты образуется рацемическая смесь, не обладающая оптической активностью.
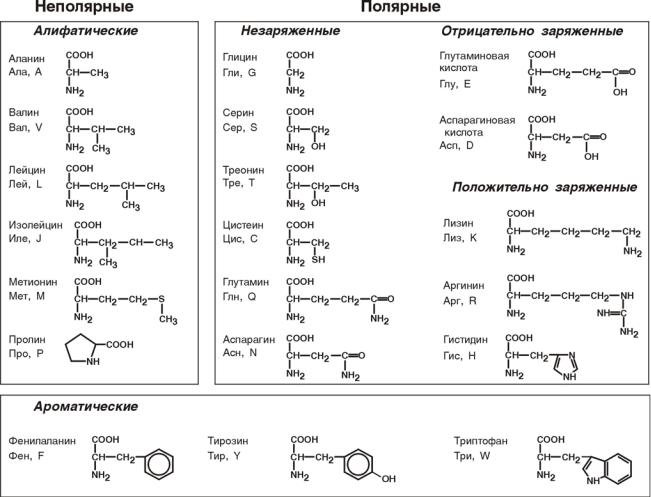
4. Для медиков наиболее интересна классификация по участию аминокислот в синтезе белков: протеиногенные (20 АК) и непротеиногенные (около 40 АК).
Протеиногенные аминокислоты подразделяют:
По строению бокового радикала – неполярные (алифатические, ароматические) и полярные (незаряженные, отрицательно и положительно заряжен-
По кислотно-основным свойствам – электрохимическая. Подразделяют нейтральные (большинство), кислые (Асп, Глу) и основные (Лиз, Арг, Гис)
аминокислоты.
По необходимости для организма (физиологическая классификация) – незаменимые (Лей, Иле, Вал, Фен, Три, Тре, Лиз, Мет) и заменимые. Две аминокислоты являются условно незаменимыми (Арг. Гис)
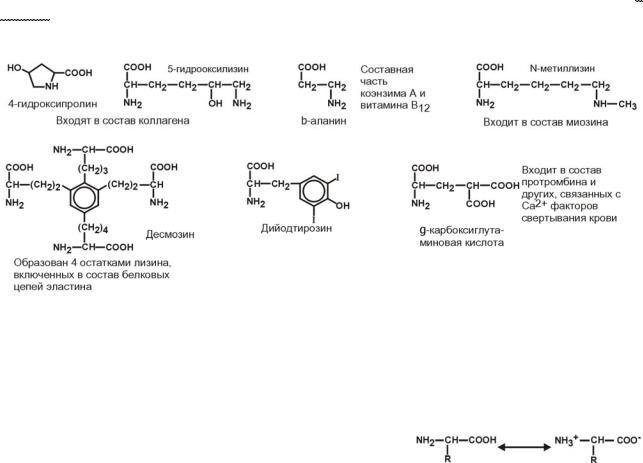
Кроме указанных, существуют и другие аминокислоты, обнаруживаемые в со-
ставе белков. В процессе синтеза белков они не участвуют, а синтезируются из про-
теиногенных аминокислот, уже находящихся в составе белковой цепи. Это нестан - дартные аминокислоты:
ФИЗИКОХИМИЧЕСКИЕ СВОЙСТВА АМИНОКИСЛОТ
1. Являются амфотерными электролитами, Аминокислоты сочетают в себе свойства и
кислот и оснований. Соответственно, в водном
растворе аминокислоты ведут себя как кислоты – доноры протонов и как основания – акцепторы
протонов.
Если общий заряд аминокислоты равен 0, то это ее состояние называют изоэлектрическим. Величина рН, при которой заряд аминокислоты равен 0, называется
изоэлектрической точкой (pI).
РI большинства аминокислот располагается в диапазоне рН от 5,5 (Фен) до
2. Заряд аминокислот зависит от величины рН среды.
Отправным пунктом для понимания причин появления заряда у аминокислот
является величина изоэлектрической точки. Ситуация различается для нейтральных, кислых и основных аминокислот.
АЗБУКА ЖИВОЙ МАТЕРИИ. БЕЛКИ Более 4 млрд лет назад на Земле из маленьких неорганических молекул непостижимым образом возникли белки, ставшие строительными блоками живых организмов. Своим бесконечным разнообразием всё живое обязано именно уникальным молекулам белка, и иные формы жизни во Вселенной науке пока неизвестны. Белки, или протеины (от греч. «протос» - «первый»), - это природные органические соединения, которые обеспечивают все жизненные процессы любого организма. Из белков построены хрусталик глаза и паутина, панцирь черепахи и ядовитые вещества грибов... С помощью белков мы перевариваем пищу и боремся с болезнями. Благодаря особым белкам по ночам светятся светлячки, а в глубинах океана мерцают таинственным светом медузы. Белковых молекул в живой клетке во много раз больше, чем всех других (кроме воды, разумеется!). Учёные выяснили, что у большинства организмов белки составляют более половины их сухой массы. И разнообразие видов белков очень велико - в одной клетке такого маленького организма, как бактерия Escherichia сой" (см. дополнительный очерк «Объект исследования - прокариоты»), насчитывается около 3 тыс. различных белков. Впервые белок был выделен (в виде клейковины) в 1728 г. итальянцем Якопо Бартоломео Беккари (1682- 1766) из пшеничной муки. Это событие принято считать рождением химии белка. С тех пор почти за три столетия из природных источников получены тысячи различных белков и исследованы их свойства. БИОЛОГИЧЕСКИЕ «БУСЫ» Молекула белка очень длинная. Химики называют такие молекулы полимерными (от греч. «поли» - «много» и «мерос» - «часть», «доля»). Действительно, длинная молекула полимера состоит из множества маленьких молекул, связанных друг с другом. Так нанизываются на нить бусинки в ожерелье. В полимерах роль нити играют химические связи между бусинками-молекулами. Секрет белков спрятан в особенностях этих самых бусинок. Большинство полимеров не принимает устойчивой формы в пространстве, уподобляясь тем же бусам, у которых и не может быть пространственной структуры: повесишь их на шею - они примут форму кольца или овала, положишь в коробку - свернутся в клубок неопределённой формы. А теперь представим себе, что некоторые бусинки могут «слипаться» друг с другом. Например, красные притягиваются к жёлтым. Тогда вся цепочка примет определённую форму, обязанную своим существованием «слипа-нию» жёлтых и красных бусинок Нечто подобное происходит и в белках. Отдельные маленькие молекулы, входящие в состав белка, обладают способностью «слипаться», так как между ними действуют силы притяжения. В результате у любой белковой цепи есть характерная только для неё пространственная структура. Именно она определяет чудесные свойства белков. Без такой структуры они не могли бы выполнять те функции, которые осуществляют в живой клетке. При длительном кипячении белков в присутствии сильных кислот или щелочей белковые цепи распадаются на составляющие их молекулы, называемые аминокислотами. Аминокислоты - это и есть те «бусинки», из которых состоит белок, и устроены они сравнительно просто. КАК УСТРОЕНА АМИНОКИСЛОТА В каждой молекуле аминокислоты есть атом углерода, связанный с четырьмя заместителями. Один из них - атом водорода, второй - карбоксильная группа -СООН. Она легко «отпускает на волю» ион водорода Н+, благодаря чему в названии аминокислот и присутствует слово «кислота». Третий заместитель - аминогруппа -NH2 и, наконец, четвёртый заместитель - группа атомов, которую в общем случае обозначают R. У всех аминокислот R-группы разные, и каждая из них играет свою, очень важную роль. Свойства «бусинок», отличающие одну аминокислоту от другой, скрыты в R- группах (их ещё называют боковыми цепями). Что же касается группы -СООН, то химики-органики относятся к ней с большим почтением: всем другим атомам углерода в молекуле даются обозначения в зависимости от степени их удалённости от карбоксильной группы. Ближайший к ней атом именуют а-атомом, второй - в-атомом, следующий - у-атомом и т. д. Атом углерода в аминокислотах, который находится ближе всех к карбоксильной группе, т. е. а- атом, связан также с аминогруппой, поэтому природные аминокислоты, входящие в состав белка, называют а-аминокислотами. В природе встречаются также аминокислоты, в которых NH^-группа связана с более отдалёнными от карбоксильной группы атомами углерода. Однако для построения белков природа выбрала именно а-аминокислоты. Это обусловлено прежде всего тем, что только а-аминокислоты, соединённые в длинные цепи, способны обеспечить достаточную прочность и устойчивость структуры больших белковых молекул. Число а-аминокислот, различающихся R-группой, велико. Но чаще других в белках встречается всего 20 разных аминокислот. Их можно рассматривать как алфавит «языка» белковой молекулы. Химики называют эти главные аминокислоты стандартными, основными или нормальными. Условно основные аминокислоты делят на четыре класса. В первый входят аминокислоты с неполярными боковыми цепями. Во второй - аминокислоты, содержащие полярную группу. Следующие два составляют аминокислоты с боковыми цепями, которые могут заряжаться положительно (они объединяются в третий класс) или отрицательно (четвёртый). Например, диссоциация карбоксильной группы даёт анион - СОО-, а протонирование атома азота - катион, например -NH3+. Боковые цепи аспарагиновой и глута-миновой кислот имеют ещё по одной карбоксильной группе -СООН, которая при значениях рН, характерных для живой клетки (рН = 7), расстаётся с ионом водорода (Н+) и приобретает отрицательный заряд. Боковые цепи аминокислот лизина, аргинина и гистидина заряжены положительно, поскольку у них есть атомы азота, которые, наоборот, могут ион водорода присоединять. Каждая а-аминокислота (кроме глицина) в зависимости от взаимного расположения четырёх заместителей может существовать в двух формах. Они отличаются друг от друга, как предмет от своего зеркального отражения или как правая рука от левой. Такие соединения получили название хоральных (от грен. «хир» - «рука»). Хиральные молекулы открыл в 1848 г. великий французский учёный Луи Пастер. Два типа оптических изомеров органических молекул получили названия Д-форма (от лат. dexter - «правый») и Z-форма (от лат. laevus - «левый»). Кстати, одно из названий других хиральных молекул - глюкозы и фруктозы - декстроза и левулоза. Примечательно, что в состав белков входят только Z-аминокислоты, и вся белковая жизнь на Земле - «левая». Для нормальной жизнедеятельности организм нуждается в полном наборе из 20 основных a-Z-аминокислот. Но одни из них могут быть синтезированы в клетках самого организма, а другие - должны поступать в готовом виде из пищевых продуктов. В первом случае аминокислоты называют заменимыми, а во втором - незаменимыми. Набор последних для разных организмов различен. Например, для белой крысы незаменимыми являются 10 аминокислот, а для молочнокислых бактерий - 16. Растения могут самостоятельно синтезировать самые разнообразные аминокислоты, создавать такие, которые не встречаются в белках. Для удобства 20 главных аминокислот обозначают символами, используя одну или первые три буквы русского или английского названия аминокислоты, например аланин - Ала или А, глицин - Гли или G. ЧТО ТАКОЕ ПЕПТИД Полимерная молекула белка образуется при соединении в длинную цепочку бусинок-аминокислот. Они нанизываются на нить химических связей благодаря имеющимся у всех аминокислот амино- и карбоксильной группам, присоединённым к а-атому углерода. Образующиеся в результате такой реакции соединения называются пеп-тидами; (-СО-NH-группировка в них - это пептидная группа, а связь между атомами углерода и азота - пептидная связь (её ещё называют амидной). Соединяя аминокислоты посредством пептидных связей, можно получить пептиды, состоящие из остатков очень многих аминокислот. Такие соединения получили название полипептиды. Полипептидное строение белковой молекулы доказал в 1902 г. немецкий химик Эмиль Герман Фишер. На концах аминокислотной цепочки находятся свободные амино-и карбоксильная группы; эти концы цепочки называют N- и С-концами. Аминокислотные остатки в полипеп-тидной цепочке принято нумеровать с N- конца. Общее число аминокислотных остатков в белковой молекуле изменяется в очень широких пределах. Так, человеческий инсулин состоит из 51 аминокислотного остатка, а лизо-цим молока кормящей матери - из 130. В гемоглобине человека 4 аминокислотные цепочки, каждая из которых построена из примерно 140 аминокислот. Существуют белки, имеющие почти 3 тыс. аминокислотных остатков в единой цепи. Молекулярные массы белков лежат в диапазоне примерно от 11 тыс. для малых белков, состоящих из 100 аминокислотных остатков, до 1 млн и более для белков с очень длинными полипептидными цепями или для белков, состоящих из нескольких по-липептидных цепей. Возникает вопрос: как же всё огромное многообразие белков с различными функциями и свойствами может быть создано всего из 20 молекул? А разгадка этого секрета природы проста - каждый белок имеет свой неповторимый аминокислотный состав и уникальный порядок соединения аминокислот, называемый первичной структурой белка. СПИРАЛИ И СЛОИ В начале 50-х гг. XX в. американские химики Лайнус Карл Полинг (1901- 1994), награждённый Нобелевской премией за исследования природы химической связи, и Роберт Кори (1897-1971) предположили, что некоторые участки аминокислотной цепочки в белках закручены в спираль. Благодаря совершенствованию экспериментальных методов (структуру белков изучают с помощью рентгеновских лучей) через несколько лет эта гениальная догадка подтвердилась. Действительно, полипептидные цепи очень часто образуют спираль, закрученную в правую сторону. Это первый, самый низкий уровень пространственной организации белковых цепочек Здесь-то и начинают играть роль слабые взаимодействия «бусинок»-аминокислот: группа С=0 и группа N-H из разных пептидных связей могут образовывать между собой водородную связь. Оказалось, что в открытой Полингом и Кори спирали такая связь образована между группой С=0 каждой г-й аминокислоты и группой N-H (i + 4)-й аминокислоты, т. е. между собой связаны аминокислотные остатки, отстоящие друг от друга на четыре «бусинки». Эти водородные связи и стабилизируют такую спираль в целом. Она получила название a.-спирали. Позднее выснилось, что а-спираль - не единственный способ укладки аминокислотных цепочек. Помимо спиралей они образуют ещё и слои. Благодаря всё тем же водородным связям между группами С=0 и N-H друг с другом могут «слипаться» сразу несколько разных фрагментов одной полипептидной цепи. В результате получается целый слой - его назвали ^-слоем. В большинстве белков а-спирали и р-слои перемежаются всевозможными изгибами и фрагментами цепи без какой-либо определённой структуры. Когда имеют дело с пространственной структурой отдельных участков белка, говорят о вторичной структуре белковой молекулы. БЕЛОК В ПРОСТРАНСТВЕ Для того чтобы получить полный «портрет» молекулы белка, знания первичной и вторичной структуры недостаточно. Эти сведения ещё не дают представления ни об объёме, ни о форме молекулы, ни тем более о расположении участков цепи по отношению друг к другу. А ведь все спирали и слои каким-то образом размещены в пространстве. Общая пространственная структура поли-пептидной цепи называется третичной структурой белка. Первые пространственные модели молекул белка - миоглобина и гемоглобина - построили в конце 50-х гг. XX в. английские биохимики Джон Ко-удери Кендрю (родился в 1917 г.) и Макс Фердинанд Перуц (родился в 1914 г.). При этом они использовали данные экспериментов с рентгеновскими лучами. За исследования в области строения белков Кендрю и Перуц в 1962 г. были удостоены Нобелевской премии. А в конце столетия была определена третичная структура уже нескольких тысяч белков. При образовании третичной структуры белка наконец-то проявляют активность R-группы - боковые цепи аминокислот. Именно благодаря им «слипаются» между собой большинство «бусинок»-аминокислот, придавая цепи определённую форму в пространстве. В живом организме белки всегда находятся в водной среде. А самое большое число основных аминокислот - восемь - содержат неполярные R-группы. Разумеется, белок стремится надёжно спрятать внутрь своей молекулы неполярные боковые цепи, чтобы ограничить их контакт с водой. Учёные называют это возникновением гидрофобных взаимодействий (см. статью «Мельчайшая единица живого»). Благодаря гидрофобным взаимодействиям вся полипептидная цепочка принимает определённую форму в пространстве, т. е. образует третичную структуру. В молекуле белка действуют и другие силы. Часть боковых цепей основных аминокислот заряжена отрицательно, а часть - положительно. Так как отрицательные заряды притягиваются к положительным, соответствующие «бусинки» «слипаются». Электростатические взаимодействия, или, как их называют иначе, солевые мостики, - ещё одна важная сила, стабилизирующая третичную структуру. У семи основных аминокислот есть полярные боковые цепи. Между ними могут возникать водородные связи, тоже играющие немалую роль в поддержании пространственной структуры белка. Между двумя аминокислотными остатками цистеина иногда образуются ковалентные связи (-S-S-), которые очень прочно фиксируют расположение разных участков белковой цепи по отношению друг к другу. Такие связи называют дисуль-фидными мостиками. Это самые немногочисленные взаимодействия в белках (в некоторых случаях они вообще отсутствуют), зато по прочности они не имеют равных. ВЫСШИЙ УРОВЕНЬ ПРОСТРАНСТВЕННОЙ ОРГАНИЗАЦИИ БЕЛКОВ Молекула белка может состоять не из одной, а из нескольких полипептидных цепей. Каждая такая цепь представляет собой самостоятельную пространственную структуру - субь-единицу. Например, белок гемоглобин состоит из четырёх субъединиц, которые образуют единую молекулу, располагаясь в вершинах почти правильного тетраэдра. Субъединицы «прилипают» друг к другу благодаря тем же самым силам, что стабилизируют третичную структуру. Это гидрофобные взаимодействия, солевые мостики и водородные связи. Если белок состоит из нескольких субъединиц, говорят, что он обладает четвертичной структурой. Такая структура представляет собой высший уровень организации белковой молекулы. В отличие от первых трёх уровней четвертичная структура есть далеко не у всех белков. Приблизительно половина из известных на сегодняшний день белков её не имеют. ПОЧЕМУ БЕЛКИ БОЯТСЯ ТЕПЛА Связи, поддерживающие пространственную структуру белка, довольно легко разрушаются. Мы с детства знаем, что при варке яиц прозрачный яичный белок превращается в упругую белую массу, а молоко при скисании загустевает. Происходит это из-за разрушения пространственной структуры белков альбумина в яичном белке и казеина (огглат. caseus - «сыр») в молоке. Такой процесс называется денатурацией. В первом случае её вызывает нагревание, а во втором - значительное увеличение кислотности (в результате жизнедеятельности обитающих в молоке бактерий). При денатурации белок теряет способность выполнять присущие ему в организме функции (отсюда и название процесса: от лат. denaturare - «лишать природных свойств»). Денатурированные белки легче усваиваются организмом, поэтому одной из целей термической обработки пищевых продуктов является денатурация белков. ЗАЧЕМ НУЖНА ПРОСТРАНСТВЕННАЯ СТРУКТУРА В природе почти ничего не происходит случайно. Если белок принял определённую форму в пространстве, это должно служить достижению какой-то цели. Действительно, только белок с «правильной» пространственной структурой может обладать определёнными свойствами, т. е. выполнять те функции в организме, которые ему предписаны. А делает он это с помощью всё тех же R-групп аминокислот. Оказывается, боковые цепи не только поддерживают «правильную» форму молекулы белка в пространстве. R-группы могут связывать другие органические и неорганические молекулы, принимать участие в химических реакциях, выступая, например, в роли катализатора. Часто сама пространственная организация полипептидной цепи как раз" и нужна для того, чтобы сосредоточить в определённых точках пространства необходимый для выполнения той или иной функции набор боковых цепей. Пожалуй, ни один процесс в живом организме не проходит без участия белков. В ЧЁМ СЕКРЕТ ФЕРМЕНТОВ Все химические реакции, протекающие в клетке, происходят благодаря особому классу белков - ферментам. Это белки-катализаторы. У них есть свой секрет, который позволяет им работать гораздо эффективнее других катализаторов, ускоряя реакции в миллиарды раз. Предположим, что несколько приятелей никак не могут встретиться. Но стоило одному из них пригласить друзей на день рождения, как результат не заставил себя ждать: все оказались в одном месте в назначенное время. Чтобы встреча состоялась, понадобилось подтолкнуть друзей к контакту. То же самое делает и фермент. В его молекуле есть так называемые цгнтры связывания. В них расположены привлекательные для определённого типа химических соединений (и только для них!) «уютные кресла» - R-группы, связывающие какие-то участки молекул реагирующих веществ. Например, если одна из молекул имеет неполярную группу, в центре связывания находятся гидрофобные боковые цепи. Если же в молекуле есть отрицательный заряд, его будет поджидать в молекуле фермента R-группа с положительным зарядом. В результате обе молекулы реагентов связываются с ферментом и оказываются в непосредственной близости друг от друга. Мало того, те их группы, которые должны вступить в химическую реакцию, сориентированы в пространстве нужным для реакции образом. Теперь за дело принимаются боковые цепи фермента, играющие роль катализаторов. В ферменте все «продумано» таким образом, что R-группы-катализаторы тоже расположены вблизи от места событий, которое называют активным центром. А после завершения реакции фермент «отпускает на волю» молекулы-продукты (см. статью «Ферменты - на все руки мастера»). ОТКУДА БЕРЁТСЯ ИММУНИТЕТ Белки выполняют в организме множество функций; они, например, защищают клетки от нежелательных вторжений, предохраняют их от повреждений. Специальные белки - антитела обладают способностью распознавать проникшие в клетки бактерии, вирусы, чужеродные полимерные молекулы и нейтрализовывать их. У высших позвоночных от чужеродных частиц организм защищает иммунная система. Она устроена так, что организм, в который вторглись такие «агрессоры» - антигены, начинает вырабатывать антитела. Молекула антитела прочно связывается с антигеном: у антител, как и у ферментов, тоже есть центры связывания. Боковые цепи аминокислот расположены в центрах таким образом, что антиген, попавший в эту ловушку, уже не сможет вырваться из «железных лап» антитела. После связывания с антителом враг выдворяется за пределы организма. Можно ввести в организм небольшое количество некоторых полимерных молекул, входящих в состав бактерий или вирусов-возбудителей какой-либо инфекционной болезни. В организме немедленно появятся соответствующие антитела. Теперь попавший в кровь или лимфу «настоящий» болезнетворный микроб тотчас же подвергнется атаке этих антител, и болезнь будет побеждена. Такой способ борьбы с инфекцией есть не что иное, как нелюбимая многими прививка. Благодаря ей организм приобретает иммунитет к инфекционным болезням. ДЛЯ ЧЕГО В ГЕМОГЛОБИНЕ ЖЕЛЕЗО В природе существуют белки, в которых помимо аминокислот содержатся другие химические компоненты, такие, как липиды, сахара, ионы металлов. Обычно эти компоненты играют важную роль при выполнении белком его биологической функции. Так, перенос молекул и ионов из одного органа в другой осуществляют транспортные белки плазмы крови. Белок гемоглобин (от греч. «гема» - «кровь» и лат. globus - «шар», «шарик»), содержащийся в кровяных клетках - эритроцитах (от греч. «эритрос» - «красный» и «китос» - «клетка»), доставляет кислород от лёгких к тканям. В молекуле гемоглобина есть комплекс иона железа Fe24" со сложной органической молекулой, называемый гемам. Гемоглобин состоит из четырёх белковых субъединиц, и каждая из них содержит по одному гему. В связывании кислорода в лёгких принимает участие непосредственно ион железа. Как только к нему хотя бы в одной из субъединиц присоединяется кислород, сам ион тут же чуть-чуть меняет своё расположение в молекуле белка. Движение железа «провоцирует» движение всей аминокислотной цепочки данной субъединицы, которая слегка трансформирует свою третичную структуру. Другая субъединица, ещё не присоединившая кислород, «чувствует», что произошло с соседкой. Её структура тоже начинает меняться. В итоге вторая субъединица связывает кислород легче, чем первая. Присоединение кислорода к третьей и четвёртой субъединицам происходит с ещё меньшими трудностями. Как видно, субъединицы помогают друг другу в работе. Для этого-то гемоглобину и нужна четвертичная структура. Оксид углерода СО (в просторечии угарный газ) связывается с железом в геме в сотни раз прочнее кислорода. Угарный газ смертельно опасен для человека, поскольку лишает гемоглобин возможности присоединять кислород. А ЕЩЁ БЕЛКИ... ...Служат питательными веществами. В семенах многих растений (пшеницы, кукурузы, риса и др.) содержатся пищевые белки. К ним относятся также альбумин - основной компонент яичного белка и казеин - главный белок молока. При переваривании в организме человека белковой пищи происходит гидролиз пептидных связей. Белки «разбираются» на отдельные аминокислоты, из которых организм в дальнейшем «строит» новые пептиды или использует для получения энергии. Отсюда и название: греческое слово «пептос» означает «переваренный». Интересно, что гидролизом пептидной связи управляют тоже белки - ферменты. ...Участвуют в регуляции клеточной и физиологической активности. К подобным белкам относятся многие гормоны (от греч. «гормао» - «побуждаю»), такие, как инсулин, регулирующий обмен глюкозы, и гормон роста. ...Наделяют организм способностью изменять форму и передвигаться. За это отвечают белки актин и миозин, из которых построены мышцы. ...Выполняют опорную и защитную функции, скрепляя биологические структуры и придавая им прочность. Кожа представляет собой почти чистый белок коллаген, а волосы, ногти и перья состоят из прочного нерастворимого белка кератина. ЧТО ЗАПИСАНО В ГЕНАХ Последовательность аминокислот в белках кодируется генами, которые хранятся и передаются по наследству с помощью молекул ДНК (см. статьи «Хранитель наследственной информации. ДНК» и «Экспрессия генов»). Пространственную структуру белка задаёт именно порядок расположения аминокислот. Получается, что не только первичная, но и вторичная, третичная и четвертичная структуры белков составляют содержание наследственной информации. Следовательно, и выполняемые белками функции запрограммированы генетически. Громадный перечень этих функций позволяет белкам по праву называться главными молекулами жизни. Поэтому сведения о белках и есть то бесценное сокровище, которое передаётся в природе от поколения к поколению. Интерес человека к этим органическим соединениям с каждым годом только увеличивается. Сегодня учёные уже расшифровали структуру многих белковых молекул. Они выясняют функции самых разных белков, пытаются определить взаимосвязь функций со структурой. Установление сходства и различий у белков, выполняющих аналогичные функции у разных живых организмов, позволяет глубже проникать в тайны эволюции. АМИНОКИСЛОТЫ - ПОКАЗАТЕЛИ ВОЗРАСТА D- и L-формы аминокислот обладают способностью очень медленно превращаться друг в друга. За определённый (весьма длительный) период времени чистая D- или I-форма может стать смесью равных количеств обеих форм. Такая смесь называется раиемагом, а сам процесс -раие-мизаиией. Скорость рацемизации зависит от температуры и типа аминокислоты. Данное свойство можно использовать для определения возраста ископаемых остатков организмов, а при необходимости - и живых существ. Например, в белке дентина (дентин - костная ткань зубов) 1-ас-парагиновая кислота самопроизвольно раиемизуется со скоростью 0,1 % в год. У детей в период формирования зубов в дентине содержится только 1-аспарагиновая кислота. Дентин выделяют из зуба и определяют В нём содержание 0-формы. Результаты теста достаточно точны. Так, для 97-летней женщины, возраст которой был документально засвидетельствован, тест показал возраст 99 лет. Данные исследований, выполненных на ископаемых остатках доисторических животных - слонов, дельфинов, медведей, - хорошо согласуются с результатами датирования, полученными радионуклидным методом. ЗА ЧТО СЕНГЕР ПОЛУЧИЛ НОБЕЛЕВСКИЕ ПРЕМИИ При гидролизе белков до аминокислот (разрушении пептидной связи водой) теряется информация о последовательности их соединения. Поэтому долгое время считали, что определение первичной структуры белка представляет собой совершенно безнадежную задачу. Но в 50-х гг. XX в. английский биохимик Фредерик Сенгер (родился в 1918 г.) смог расшифровать последовательность аминокислот в полипептидных цепях гормона инсулина. За эту работу, на выполнение которой ушло несколько лет, в 1958 г. Сенгер был удостоен Нобелевской премии по химии (двадцатью годами позже он совместно с У. Гилбертом получил вторую премию за вклад в установление первичной структуры ДНК). Принципы определения аминокислотной последовательности, впервые сформулированные Сенгером, используются и ныне, правда, со всевозможными вариациями и усовершенствованиями. Процедура установления первичной структуры белка сложна и многоступенчата: в ней около десятка различных стадий. Сначала белок расщепляют до отдельных аминокислот и устанавливают их тип и количество в данном веществе. На следующей стадии длинную белковую молекулу расщепляют уже не полностью, а на фрагменты. Затем в этих фрагментах определяют порядок соединения аминокислот, последовательно отделяя их одну за другой. Расшепление белка на фрагменты проводят несколькими способами, чтобы в разных фрагментах были перекрывающиеся участки. Выяснив порядок расположения аминокислот во всех фрагментах, получают полную информацию о том, как аминокислоты расположены в белке. К концу XX в. созданы специальные приборы, определяющие последовательность аминокислот в молекуле белка в автоматическом режиме - секвенаторы (от англ. sequence - «последовательность»). молоко И КИСЛОМОЛОЧНЫЕ ПРОДУКТЫ Молоко представляет собой коллоидный раствор жира в воде. Под микроскопом хорошо видно, что оно неоднородно: в бесцветном растворе (сыворотке) плавают жировые шарики. В коровьем молоке обычно содержится от 3 до 6 % жиров (в основном это сложные эфиры глицерина и насыщенных карбоновых кислот - пальмитиновой, стеариновой), около 3 % белков, а ешё углеводы, органические кислоты, витамины и минеральные вещества. Белок казеин в молоке присутствует в связанном виде - ковалентно присоединённые к аминокислоте сери-ну фосфатные группы образуют соли с ионами кальция. При подкислении молока эти соли разрушаются, и казеин выделяется в виде белой
Полнотекстовый поиск:
Главная > Реферат >Медицина, здоровье
Аминокислоты и белки
Строительными блоками белков служат аминокислоты. Классификация аминокислот.
Моноаминомонокарбоновые: Глицин, аланин, валин, лейцин, изолейцин.
Моноаминодикарбоновые: глутаминовая и аспаргиновая кислоты.
Диаминомонокарбоновые: аргинин, лизин, оксилизин.
Ароматические: фенилаланин, тирозин.
Гетероциклические: триптофан, пролин, оксипролин, гистидин.
Аминокислота представляет собой производное органической кислоты, в котором водород в α-положении замещен на аминогруппу (-NH 2). Например, из уксусной кислоты образуется глицин, а из пропионовой - аланин. В аминокислотах одновременно присутствуют и кислотная и основная группы (карбоксил -СООН и аминогруппа -NH 2), они относятся к амфотерным соединениям .
Присутствующие в клетке свободные аминокислоты образуются в результате расщепления белков или поступают из межклеточной жидкости. Свободные аминокислоты составляют так называемый аминокислотный фонд, из которого клетка черпает строительные блоки для синтеза новых белков.
Связь R-NH-СО-R называется пептидной связью. Образующаяся молекула также является амфотерной, поскольку на одном ее конце всегда находится кислая группа, а на другом - основная; боковые цепи (остатки аминокислот) могут быть основными или кислыми. Комбинация из двух аминокислот носит название дипептида, из трех - трипептида. Пептид, состоящий из небольшого числа аминокислот, называется олигопептидо.и. Если же число аминокислот в молекуле достаточно велико, вещество называют полипептидом.
Расстояние между двумя пептидными связями равно примерно 0,35 нм. Молекула белка с мол. массой 30 000, состоящая из 300 аминокислотных остатков, в полностью вытянутом состоянии должна иметь длину 100 нм, ширину 1 нм и толщину 0,46 нм.
Белки называют протеинами (греч. протео - занимаю первое место). Это слово [в русском языке оно сохранилось лишь в названиях сложных белков] указывает, что все основные функции организма связаны со специфическими белками. Они входят в состав ферментов и сократительного аппарата клеток, присутствуют в крови и других межклеточных жидкостях. Некоторые длииноцепочечные белки, такие, как коллаген и эластин, играют важную роль в построении тканевых структур.
Кератин и кол лаген нерастворимы и обладают фибриллярной структурой; глобулярные белки, например яичный альбумин и белки сыворотки, растворимы в воде и солевых растворах и их молекулы имеют сферическую, а не нитевидную форму.
Сложные белки, в молекулу которых входит небелковая часть, так называемая простетическая группа. К ним принадлежат нуклеопротеиды , липо протеиды и хромопротеиды (гемоглобин, гемоцианин и цитохромы), в которых простетической группой служит пигмент. Простетической группой гемоглобина и миоглобина (белка мышц) является гем - металлсодержащее органическое соединение, связывающее кислород.
Первичная структура белков . Полипептидная цепь, построенная из аминокислот, представляет собой первичную структуру белковой молекулы. Это наиболее важная специфическая структура, до некоторой степени определяющая так называемые вторичную и третичную структуры белка. Агрегаты белковых субъединиц, обладающих вторичной и третичной структурой, составляют четвертичную структуру.
Изучение порядка расположения аминокислот в молекуле белка стало возможным после того, как были разработаны методы расщепления белков. Первый успех принадлежит Сэнджеру, которому в 1954 г. удалось, наконец, полностью расшифровать последовательность аминокислот в инсулине. Молекула инсулина состоит из двух цепей: А-цепь содержит 21 аминокислоту, а В-цепь - 30. Обе цепи соединены двумя дисульфидными (-S-S-) связями.
В молекуле белка аминокислоты уложены как бусины на нити, и последовательность их расположения имеет важное биологическое значение. Например, ферментативные свойства некоторых белков определяются последовательностью аминокислот на небольшом участке цепи, называемом активным центром . В молекуле гемоглобина замена одной-единственной аминокислоты уже приводит к глубоким биологическим изменениям.
Вторичная структура белков . Молекула белка состоит из нескольких сотен аминокислот, и поэтому полипептидная цепь лишь в редких случаях бывает вытянута полностью; обычно она определенным образом изогнута, образуя вторичную структуру. Фибриллярные белки (склеропротеины) часто характеризуются упорядоченным расположением цепей, благодаря чему их можно исследовать методом рентгеноструктурного анализа. В результате этих исследований было найдено, что фибриллярные белки можно разбить на три структурных типа или группы.
В белках типа β-кератина смежные цепи расположены таким образом, что образуют струк туру складчатого слоя . В этой структуре боковые группы (аминокислотные остатки) перпендикулярны плоскости, в которой лежат сами цепи; отдельные цепи соединены друг с другом водородными связями, образуя «пептидную решетку».
В белках типа α-кератина полипептидная цепь закручена в виде спирали, образуя так называемую а-спиральную структуру . Водородные связи в этом случае являются внутримолекулярными, а не межмолекулярными. Для группы коллагена предложена модель, состоящая из трех спиралей.
Третичная структура белков . В глобулярных белках полипептидные цепи определенным образом свернуты, образуя компактную структуру. Расположение таких цепей в пространстве очень сложно, но может быть выяснено методом рентгеноструктурпого анализа.
Пространственное расположение цепей до некоторой степени предопределено последовательностью чередования аминокислот в первичной структуре и связями, образующимися между некоторыми аминокислотными остатками. Многие биологические свойства белков, например ферментативная активность и антигенноетъ, связаны именно с третичной структурой.
Четвертичная структура белка; прин цип самосборки. В отличие от первичной, вторичной и третичной структур, которые содержат одну полипептидную цепь, четвертичная структура состоит из двух или более цепей. Эти цепи могут быть одинаковыми или разными, но в обоих случаях они связаны слабыми связями (нековалентнымн). Например, молекула гемоглобина состоит из четырех полипептидных субъединиц - двух α и двух β-цепей. Разделение и ассоциация этих субъединиц может происходить спонтанно.Под действием мочевины молекула гемоглобина распадается на две половники, одна из которых состоит из двух α-субъединиц, в другая из двух β -субъединиц. При удалении мочевины они объединяются вновь, образуя четырехкомпонентную молекулу. Этот процесс высокоспецифичен: объединяться могут только две разные половинки молекул (так называемый принцип самосборки). Многие ферменты и другие белки с мол. массой свыше 50 000, вероятно, обладают четвертичной структурой. Например, альдолаза (мол. масса 150 000) распадается при низком рН на субъединицы с мол. массой 50 000 каждая, но вновь ассоциирует при нейтральном рН.
Связи в белковой молекуле . В структуре белков встречаются самые различные типы связей. Первичная структура (пептидная связь) полностью определяется химическими, или ковалентными , связями. Между остаткам цистина (например, в инсулине и рибонуклеазе) образуются S-S-связи той же природы. Вторичная и третичная структуры стабилизируются рядом более слабых связей. Эти связи можно классифицировать следующим образом:
Ионные, или электростатические, связи между положительными и отрицательными ионами, находящимися на расстоянии 0,2...0,3 нм.
Водородные связи (длина связи 0,25...0,32 нм); эти по существу также электростатические связи, но более слабые, чем ионные, образуются между двумя сильно отрицательными атомами - С, N или О.
Слабые связи между неполярными боковыми цепями, возникающие в результате взаимного отталкивания молекул растворителя.
Связи, образующиеся за счет вандерваальсовых сил при взаимодействии полярных боковых цепей.
Электрические заряды белков . Все аминокислоты являются амфолитами (цвиттерионами), обладающими положительно и отрицательно заряженными группами (-NH 2 и -СООН). Так как эти группы участвуют в образовании пептидной связи, в полипептидной цепи свободными остаются только концевые СООН- и - NH 2- группы, а также СООН-группы из дикарбоновых аминокислот и NH 2 -группы из диаминокислот. Все эти группы ионизируются следующим образом:
Кислые группы теряют протоны и становятся отрицательно заряженными. Этот тип диссоциации встречается в дикарбоновых аминокислотах (аспарагиновая и глутаминовая), у которых свободная карбоксильная группа диссоциирует на СОО - и Н + .
Основные группы, приобретая протон, становятся положительно заряженными. Этот тип встречается в аминокислотах с двумя основными группами (лизин и аргинин), у которых свободные аминогруппы ионизируются с образованием положительного заряда.
Все эти так называемые ионогенные группы вместе с концевыми свободными карбоксильными и аминогруппами участвуют в кислотно-щелочных реакциях белков и определяют электрические свойства белковых молекул.
Движение
белков в электирическом поле -
электрофорез.
Аминокислоты
- соединения, содержащие амино- и
карбоксильную
группы. В зависимости от расположения
амино- и карбоксильной
групп различают α-, β-, γ-,
δ-
и т. д. аминокислоты:
α-Аминокислоты являются составными частями белков и участвуют в важнейших биологических процессах. Первая аминокислота была выделена в 1820 г. французским исследователем X. Браконно кислотным гидролизом желатины, однако лишь через 13 лет в ней было обнаружено присутствие азота. Позднее была показана роль α-аминокислот как структурных элементов белка (Н. Н. Любавин, 1871 г.). К началу XX в. методом гидролиза белка было выделено более 20 аминокислот.
Для синтеза белков и других биохимических реакций организм использует исключительно аминокислоты, а не белки, поступающие с пищей. Некоторые аминокислоты, необходимые для роста и нормального функционирования животных организмов, потребляются готовыми из пищи, так как скорость их синтеза отстает от скорости расхода. Такие аминокислоты называются незаменимыми аминокислотами. К ним относятся валин, лейцин, изолейцин, фенилаланин, аргинин, треонин, метионин, лизин, триптофан, гистидии.
Для α-аминокислот характерны исторические названия. Их происхождение связано со свойствами и названиями продуктов, из которых они впервые были выделены. Глицин имеет сладкий вкус (от греч. «глюкос» - сладкий). Цистин выделен из камней желчного пузыря (от греч. «цистис» -пузырь). Лейцин получен из молочного белка - казеина (от греч. «леукос» - белый). Аспарагиновая кислота изолирована из ростков спаржи (от греч. аспарагус - спаржа). Орнитин выделен из помета птиц (от греч. «ор-нитус» - птица). Аминокислоты называют также по названиям материнских карбоновых кислот. Положение аминогруппы и других заместителей обозначают буквами греческого алфавита. Научная номенклатура к аминокислотам обычно не применяется.
Все природные аминокислоты (кроме глицина) оптически активны и принадлежат к L-ряду. При микробиологическом способе аминокислоты образуются в процессе жизнедеятельности бактерий. Гидролитический метод основан на гидролизе белковых природных продуктов, например рогов, копыт, крови (отходов преимущественно мясной промышленности), из которых выделяются аминокислоты. Оба способа приводят к получению смеси оптически активных α-аминокислот L-ряда. Синтетические методы дают рацемическую смесь D- и L-аминокислот.
В производственных масштабах микробиологическим методом преимущественно получают лизин и глутаминовую кислоту; гидролитическим - цистеин, лейцин, изолейцин; синтетическим - метионин и глутаминовую кислоту. В отдельных случаях сочетают синтетический и микробиологический способы (лизин). Сначала синтезируют рацемическую смесь аминокислот, а затем ферментативно, в результате поглощения бактериями D-изомера, выделяют L- изомер.
Недавно α-аминокислоты получали в незначительных количествах и использовали преимущественно для научных исследований. Сейчас они стали многотоннажными промышленными продуктами в связи с необходимостью обеспечения питанием все возрастающего населения земного шара, из которого, по меньшей мере голодают 500 млн. и недоедает 1 млрд. человек.
Неполноценность пищи заключается преимущественно в нехватке белков, которые в желудочно-кишечном тракте гидролизуются до аминокислот L-ряда. Наибольшее значение имеют незаменимые пищевые кислоты: L-лизин, L-триптофаи, L-метионин и L-глутаминовая кислота. Белковое голодание определяется сейчас в 4 млп. т белка, соответствующих 15 млн. т мяса крупного рогатого скота. Оно преодолевается увеличением ресурсов сельского хозяйства (животноводство и земледелие), получением из углеводородов нефти микробиологического белка (кормового и пищевого) без вкуса и запаха, не уступающего по питательности пищевым белкам, богатым лизином, но лишенным метионина. Наконец, питательная ценность пищи и кормов значительно повышается добавлением к ним небольших количеств незаменимых α-аминокислот. Так, например, добавление 0,1-0,25% лизина к кормам снижает расход кормов на 15-20% и увеличивает привес сельскохозяйственных животных на 20%, а введение в корм метионина повышает яйценосность кур на 20%. Глутаминова я_кислота - самая распространенная в мире приправа (после соли)-добавляется для улучшения вкуса почти всех пищевых концентратов и консервов. Она также помогает бороться с некоторыми нервно-психическими заболеваниями.
α-Аминокислоты являются основным компонентом синтетической пищи на углеводной основе, содержащей необходимые витамины и синтетические вкусовые вещества. Поэтому возросшая потребность в α-аминокислотах потребовала разработки простых и дешевых промышленных способов их получения с использованием доступных исходных продуктов.
Синтез аминокислот. Аминирование α-галогенокарбоновых кислот - первый синтетический метод получения аминокислот (У. Перкин, 1858 г.).
Физические и химические свойства. α-Аминокислоты - твердые кристаллические вещества, вследствие ионного строения имеют высокие и нечеткие температуры плавления, обычно хорошо растворимы в воде, плохо в спирте и совсем не растворяются в эфире.
Некоторые β-замещенные γ-аминомасляной кислоты (β-фенил-γ-аминомасляная кислота, фенибут - по официальной терминологии) являются психотропными успокаивающими медицинскими препаратами (транквилизаторами), благоприятствующими улучшению состояния психических больных и снижающими нервное напряжение у здоровых людей. Их основным преимуществом по сравнению с многочисленными психотропными препаратами, чуждыми организму по химическому строению, является отсутствие токсичности вследствие близости строения к естественным продуктам обмена. Воздействие лекарственных веществ - продуктов синтетической органической химии - на психику изучает новая ветвь фармакологии - психофармакология.
Растения и большая часть микроорганизмов способны производить весь набор аминокислот и, следовательно, располагают набором всех ферментов, необходимых для их биосинтеза. У животных, аналогично тому, как это имеет место в случае коферментов и кофакторов, часть ферментов, необходимых для биосинтеза аминокислот из простых и доступных предшественников, отсутствует, в связи с чем некоторые аминокислоты должны быть получены ими с пищей. Такие аминокислоты называют незаменимыми. К их числу относят триптофан, фенилалапин, валин, изолейцин, лейцин, метионин, лизин, аргинин, гнетидин и треонин. Строго говоря, к этой же категории следовало бы отнести цистепп и тирозин, поскольку пути их биосинтеза у этих организмов из доступных предшественников отсутствуют. Однако в продуктах питания их присутствие не столь обязательно, так как цистеин может легко образовываться из незаменимого метионини, а тирозин - из незаменимого фенил-аланина. Аргинин является незаменимой аминокислотой лишь в период интенсивного роста организмов, когда он необходим в особенно больших количествах. Умеренные потребности в аргинине у животных могут обеспечиваться за счет функционирования цикла мочевины.
Полностью заменимыми являются восемь аминокислот: аланин, аспартат, аспарагин, глутамат, глутамин, серин, глицин и пролин.
Глицин используется в качестве строительного блока при синтезе пуриновых колец.
Выполняют функцию биокатализаторов - ферментов, регулирующих...


